















 {{item.name}}会员
{{item.name}}会员

Nature子刊 | 线粒体与糖酵解协同作用中HK1的O-GlcNAc糖基化修饰的调控机制
Highlights
1. HK1的O-GlcNAcylation对其在线粒体的定位和细胞代谢效率有重要影响。
2. OGT通过O-GlcNAcylation促进HK1和其他糖酵解酶在线粒体上的聚集,形成代谢体,提高能量产生。
近日,“Nature Metabolism”(IF=18.9)上发表了一篇题为“Organization of a functional glycolytic metabolon on mitochondria for metabolic efficiency”的文章。这篇文章研究了糖酵解过程中的关键限速酶己糖激酶1(HK1)在能量需求高的组织中的活性调控机制。

研究背景介绍
己糖激酶1(HK1)是糖酵解过程中的首个限速酶,负责将葡萄糖磷酸化为葡萄糖-6-磷酸。HK1在能量需求高的细胞中尤为重要,如神经元,其活性和线粒体的关联对细胞能量代谢至关重要。
O-乙酰葡糖胺糖基化修饰(O-GlcNAcylation)是一种翻译后修饰,通过将 O-乙酰葡萄糖胺添加到蛋白质丝氨酸和苏氨酸残基上来调节蛋白质功能。其水平受葡萄糖代谢和细胞能量状态的影响,是细胞营养感应和代谢调节的关键机制。
O-GlcNAc转移酶(OGT)是一种代谢传感器酶,负责催化蛋白质的O-GlcNAcylation。OGT活性受葡萄糖流量的影响,通过调节O-GlcNAcylation影响蛋白质功能和细胞代谢。
研究思路分析

研究技术路线图
01HK1的定位与调控
①免疫荧光分析发现HK1在大鼠海马神经元中与线粒体共定位在细胞体、树突和轴突中。对神经元进行了葡萄糖浓度变化的实验,包括降低葡萄糖浓度后再恢复,观察到HK1在线粒体上的定位随葡萄糖浓度的变化而变化。
②对小鼠进行喂食和禁食实验,发现血液葡萄糖水平的波动与喂食周期相符。在检查海马CA3区HK1定位时,发现与血液葡萄糖水平有明确相关性。禁食导致的血糖下降会使线粒体上的HK1表达下调,而重新喂食后血糖上升则会导致线粒体HK1表达上调。而喂食或禁食并未改变HK1的整体表达水平。
③研究中,己糖胺生物合成途径产生的UDP-GlcNAc是O-GlcNAcylation的一个关键底物。该途径通过OGT动态调节O-GlcNAc的添加,并通过O-GlcNAcase(OGA)的酶活性来平衡其移除。通过药理和遗传操作这两种酶并调节O-GlcNAcylation水平,发现增强或减少O-GlcNAcylation均会增加或减少线粒体中的HK1。此外,OGT抑制剂的存在减弱了葡萄糖通量依赖的HK1在线粒体上的招募。且线粒体大小不受O-GlcNAcylation水平波动的影响。
02O-GlcNAcylation对HK1功能的影响
①通过基因工程,使秀丽隐杆线虫的DA9神经元内源性地表达带有7xSplitGFP标签的HK1。使用OGT抑制剂处理线虫使其不过度表达OGA的情况下降低O-GlcNAcylation,观察到O-GlcNAcylation的减少与线粒体HK1的减少相对应。
②利用CRISPR技术在HEK293T细胞中对内源性HK1进行标记,分析发现OGT活性的增加和OGA活性的抑制会导致HK1的O-GlcNAcylation水平上升。此外,这种修饰在HK2上的水平则不受OGT和OGA活性变化的影响。
③通过在COS-7细胞里共表达GFP标记的HK1和分裂的OGA(nGFP–OGA)来选择性地降低HK1的O-GlcNAcylation。当同时表达myc–OGA(1–400)和nGFP–HA–OGA(544–706)时,OGA的酶活性才存在。通过固定细胞并使用线粒体标记蛋白Tomm20进行染色,发现降低HK1的O-GlcNAcylation会导致COS-7细胞中线粒体HK1的减少。
④通过免疫沉淀法从HEK293T细胞中分离HK1蛋白,并在不同O-GlcNAcylation条件下进行分析,确认了HK1的O-GlcNAcylation水平在O-GlcNAcylation环境增强时上升。通过质谱分析,发现HK1上唯一的O-GlcNAc修饰位点为T259。计算模型分析显示,T259的O-GlcNAcylation导致HK1结构发生大尺度重排,失去了部分二级结构,并使得结合口袋变得松弛,这可能会影响HK1与线粒体的相互作用。
03HK1在代谢和神经功能中的作用
①通过定点突变将HK1上的Thr259替换为丙氨酸,消除了该位点的O-GlcNAcylation。在HEK293T细胞中表达野生型HK1和T259A-HK1突变体,成像实验显示,在模拟葡萄糖通量增加的条件下,野生型HK1的线粒体定位显著增加,而T259A-HK1突变体则没有这种效果。在海马神经元中表达T259A-HK1–GFP,并通过调节O-GlcNAcylation水平,发现T259位点缺乏O-GlcNAcylation使得HK1的线粒体定位对O-GlcNAcylation水平的变化不敏感,且不影响线粒体大小。
②实验发现O-GlcNAcylation水平的增加可以提高G6P水平,促进葡萄糖磷酸化,并增强糖酵解和线粒体氧化磷酸化的ATP产生率。此外,T259位点的O-GlcNAcylation缺失会导致HK1的活性受损,但通过将其靶向募集到线粒体,可以恢复其酶活性,从而提高ATP产生率。
04HK1在神经活动中的作用
①研究发现,在特定条件下,如缺氧,糖酵解酶可以形成代谢体以提高代谢效率。通过在HEK239T细胞中提高O-GlcNAcylation水平,发现这不仅增加了线粒体中的HK1,还增加了PKM2和Aldo A的水平,而减少了它们在细胞质中的量。此外,使用成像技术证实了增强的O-GlcNAcylation促进了PKM2在线粒体上的关联。表明,O-GlcNAcylation通过调节HK1的线粒体定位,促进了糖酵解酶的招募和代谢体的形成,从而提高了ATP的产生率。
②研究发现,HK1在线粒体的定位可以通过G6P浓度调节,其中高G6P浓度会导致HK1释放,作为一种负反馈机制。通过计算模拟和实验,证实了O-GlcNAcylation降低了HK1与G6P的亲和力,并减少了HK1从线粒体的释放。实验中,通过改变G6P浓度,观察到O-GlcNAcylation水平的提高减少了HK1的释放,而T259A突变型HK1则不受影响。
③通过活细胞成像技术测量了大鼠海马神经元中的轴突ATP水平和突触囊泡回收率。实验结果表明,与野生型HK1相比,T259A-HK1突变型的表达导致ATP水平无法恢复,并且突触囊泡回收率降低。通过使用河豚毒素抑制神经元活动以保存ATP,可以恢复T259A-HK1表达神经元的突触囊泡回收。此外,使用GCaMP6s钙离子指示剂的结果显示,HK1的O-GlcNAcylation对突触囊泡回收的调控作用独立于钙离子处理。
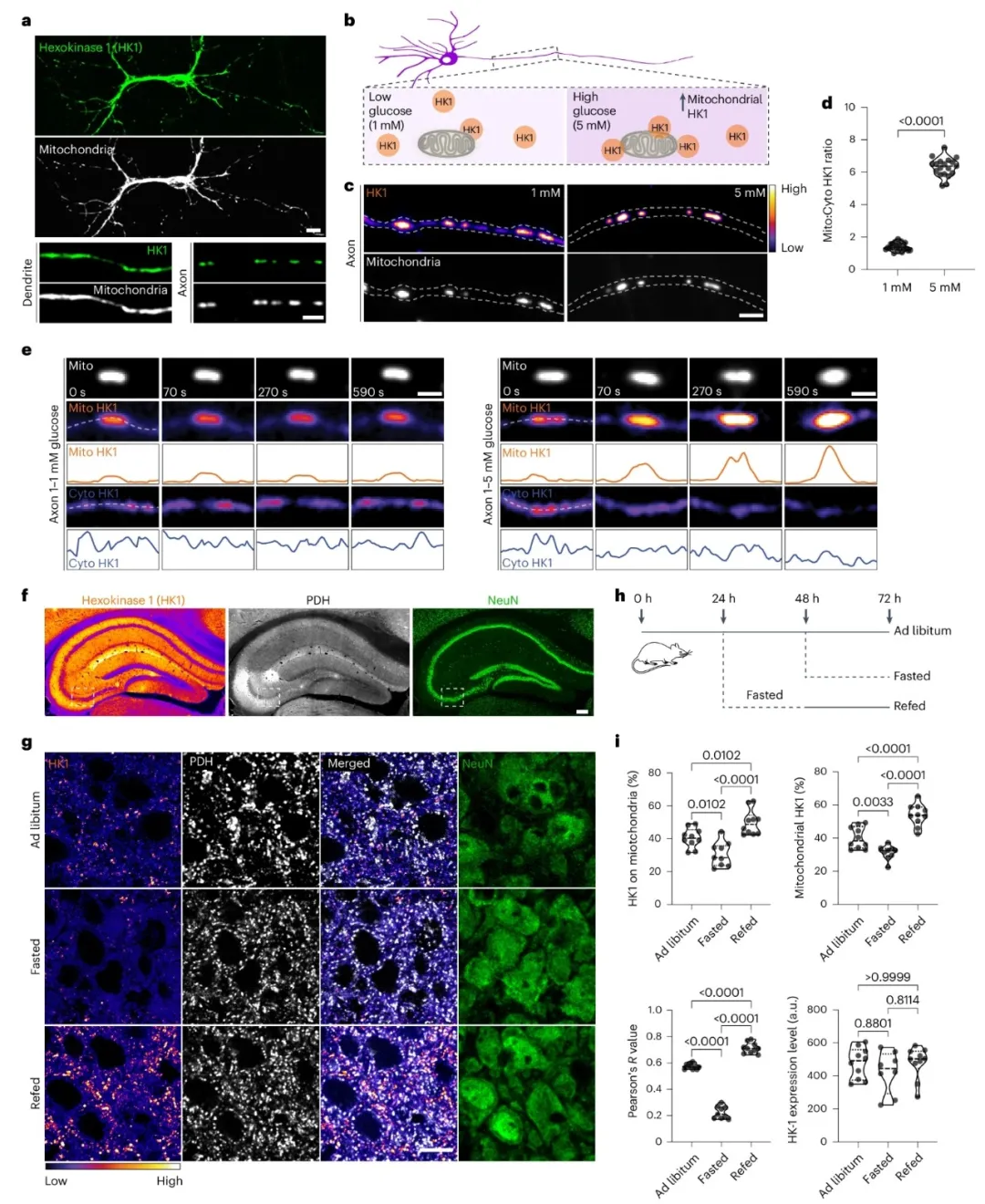
图1. HK1定位的葡萄糖依赖性调节
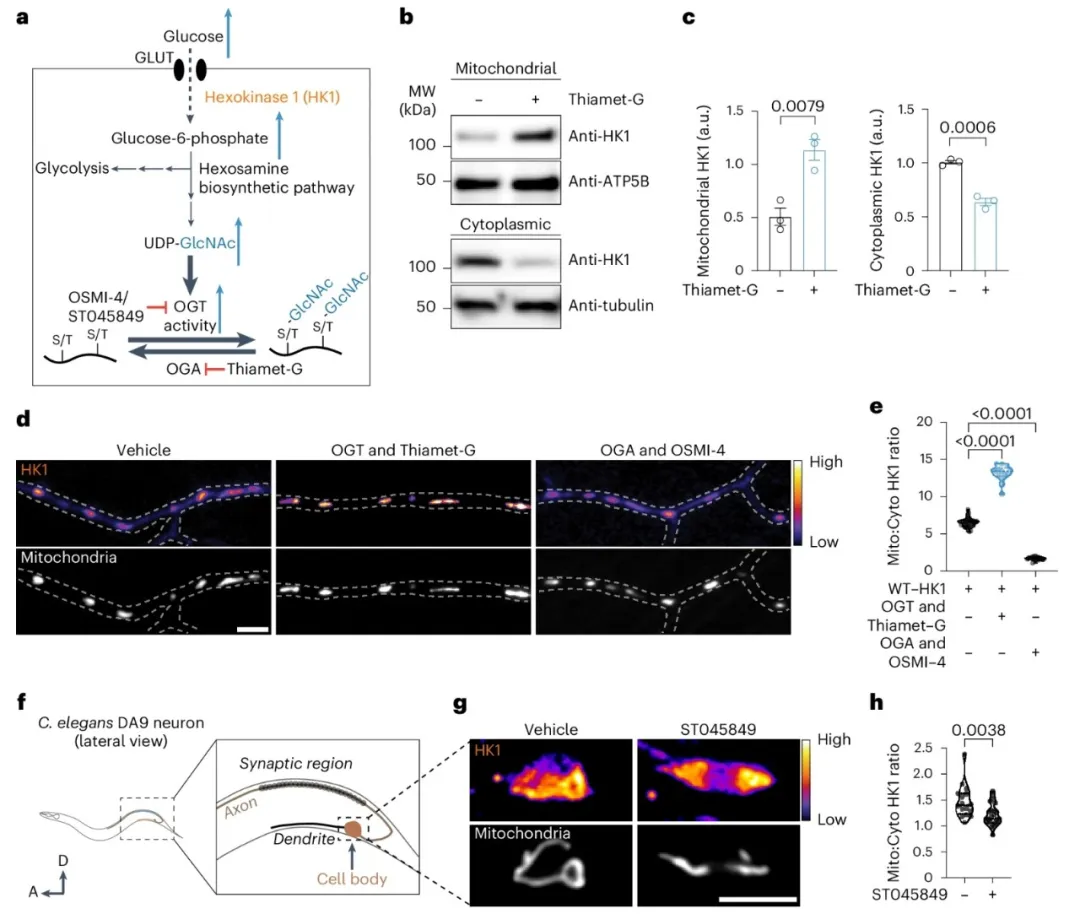
图2. O-GlcNAcylation调节HK1的线粒体定位
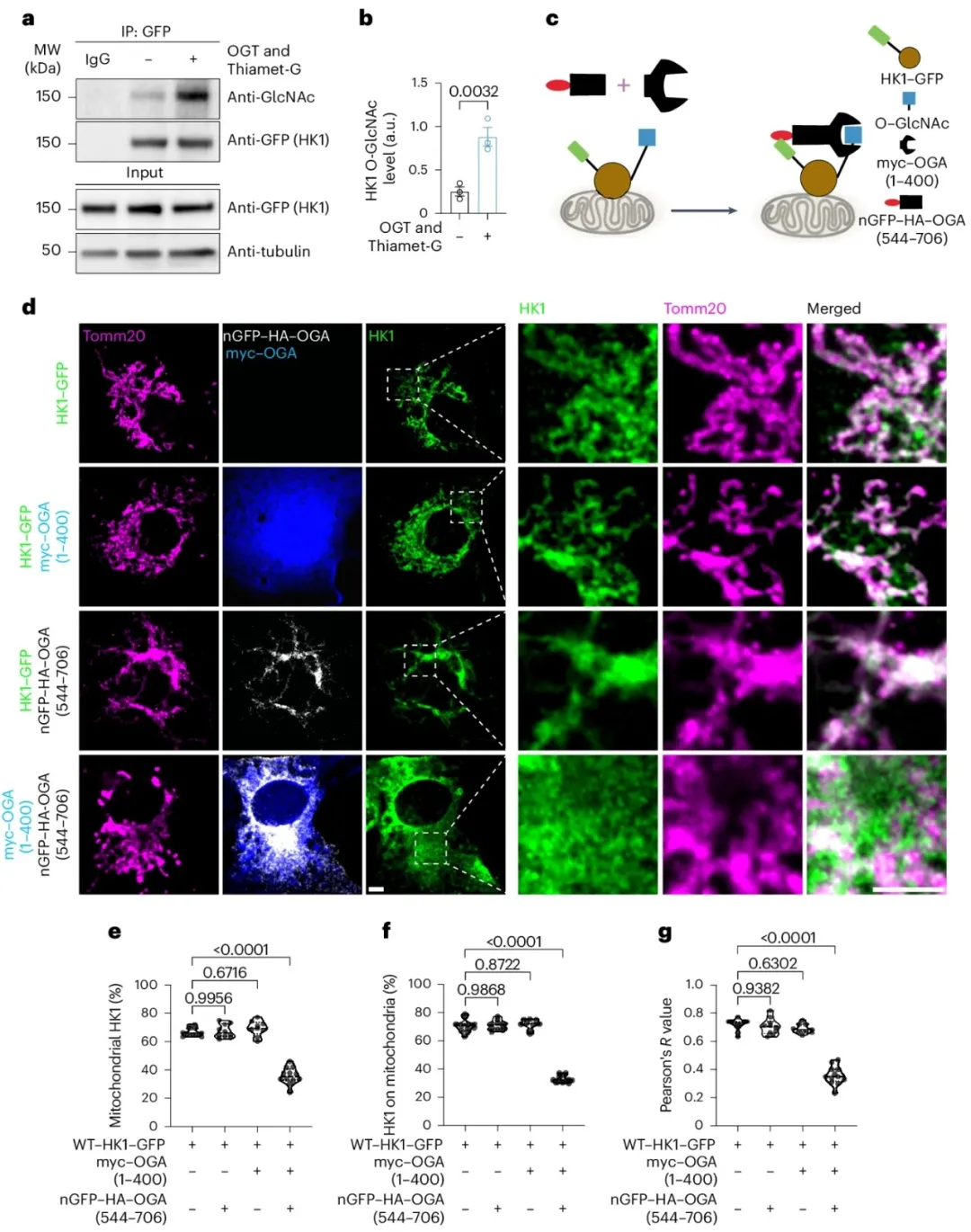
图3. OGT和OGA调节HK1 O-GlcNAcylation
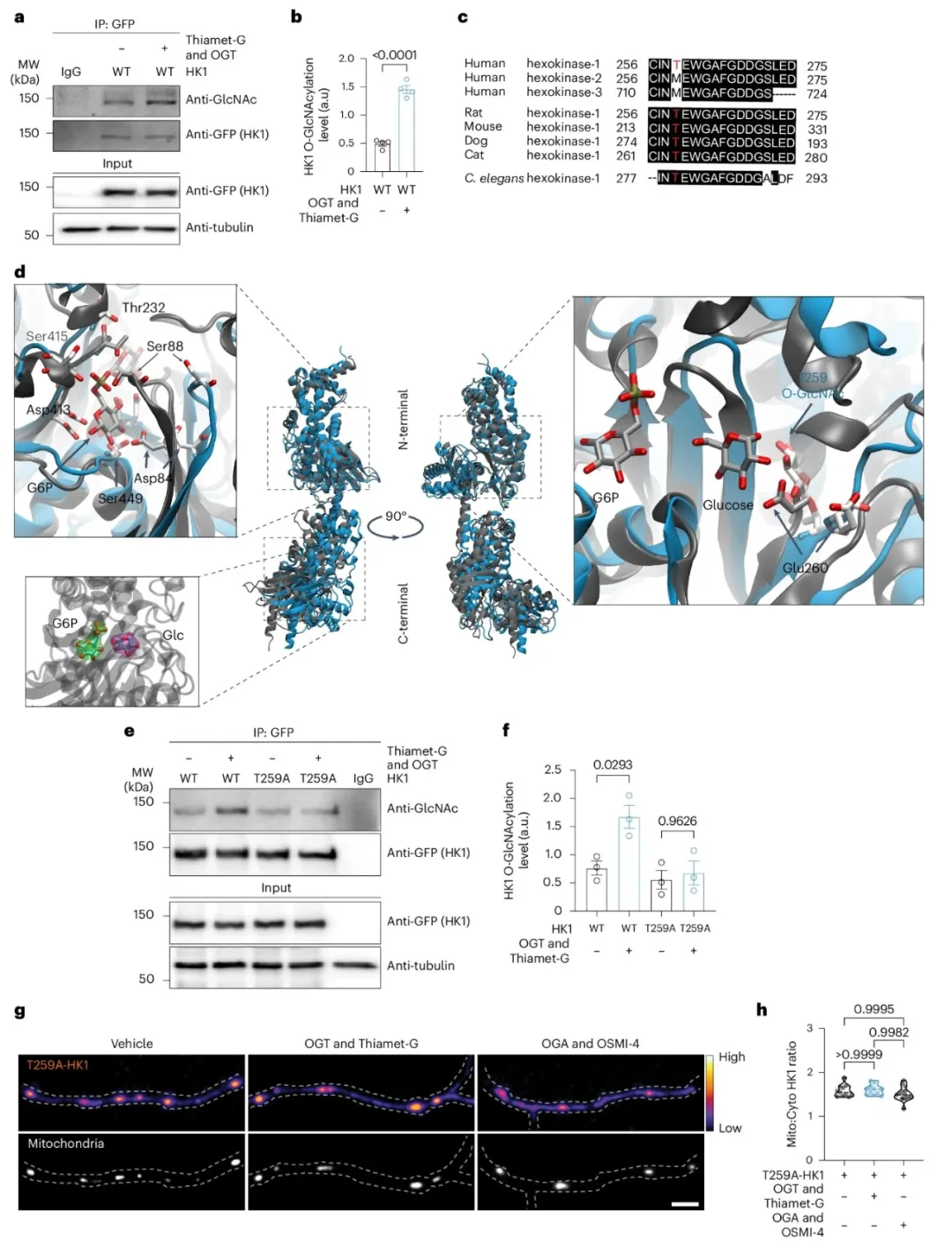
图4. HK1定位的OGT依赖性调节需要O-GlcNAcylation
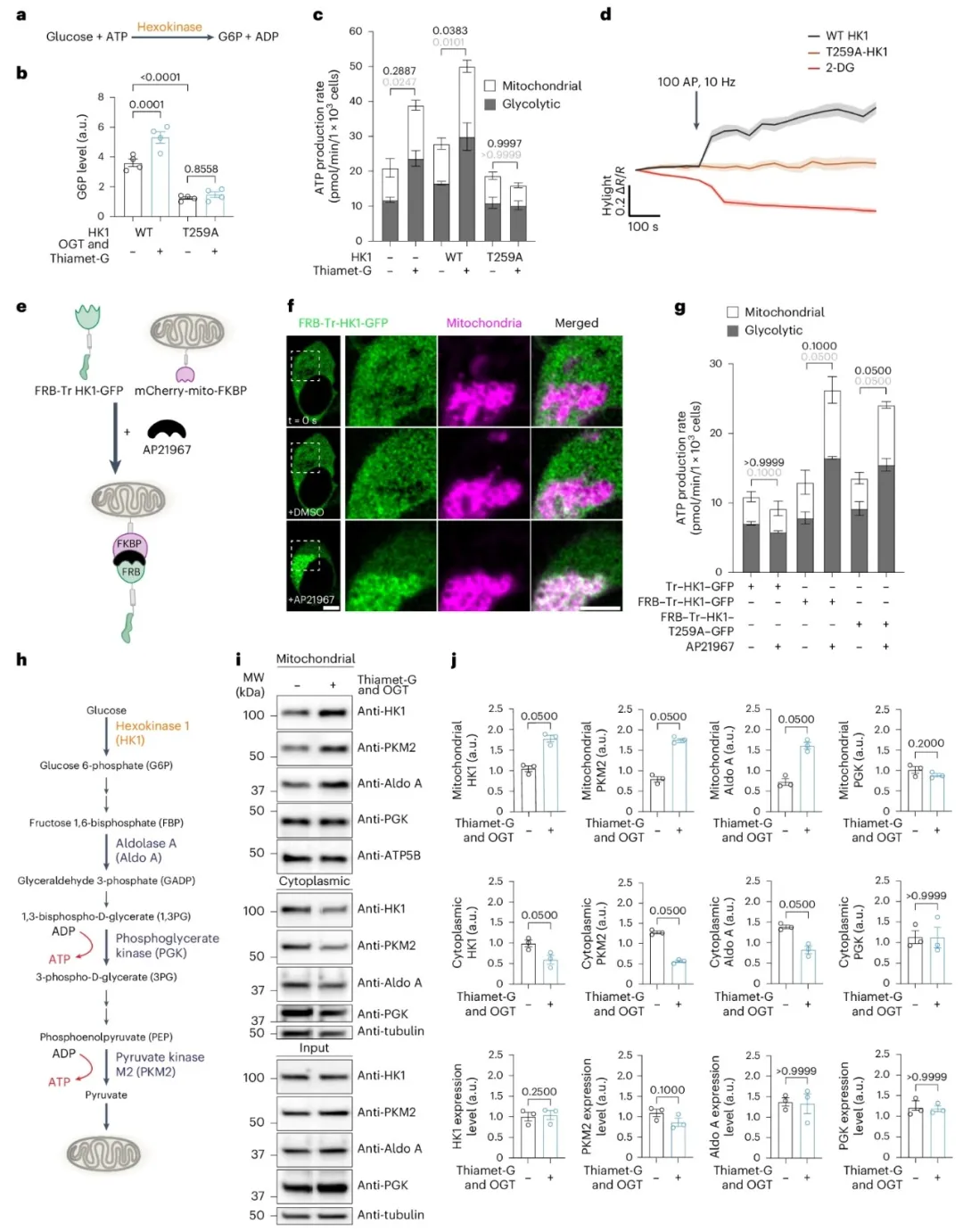
图5. HK1 O-GlcNAcylation提高了代谢效率
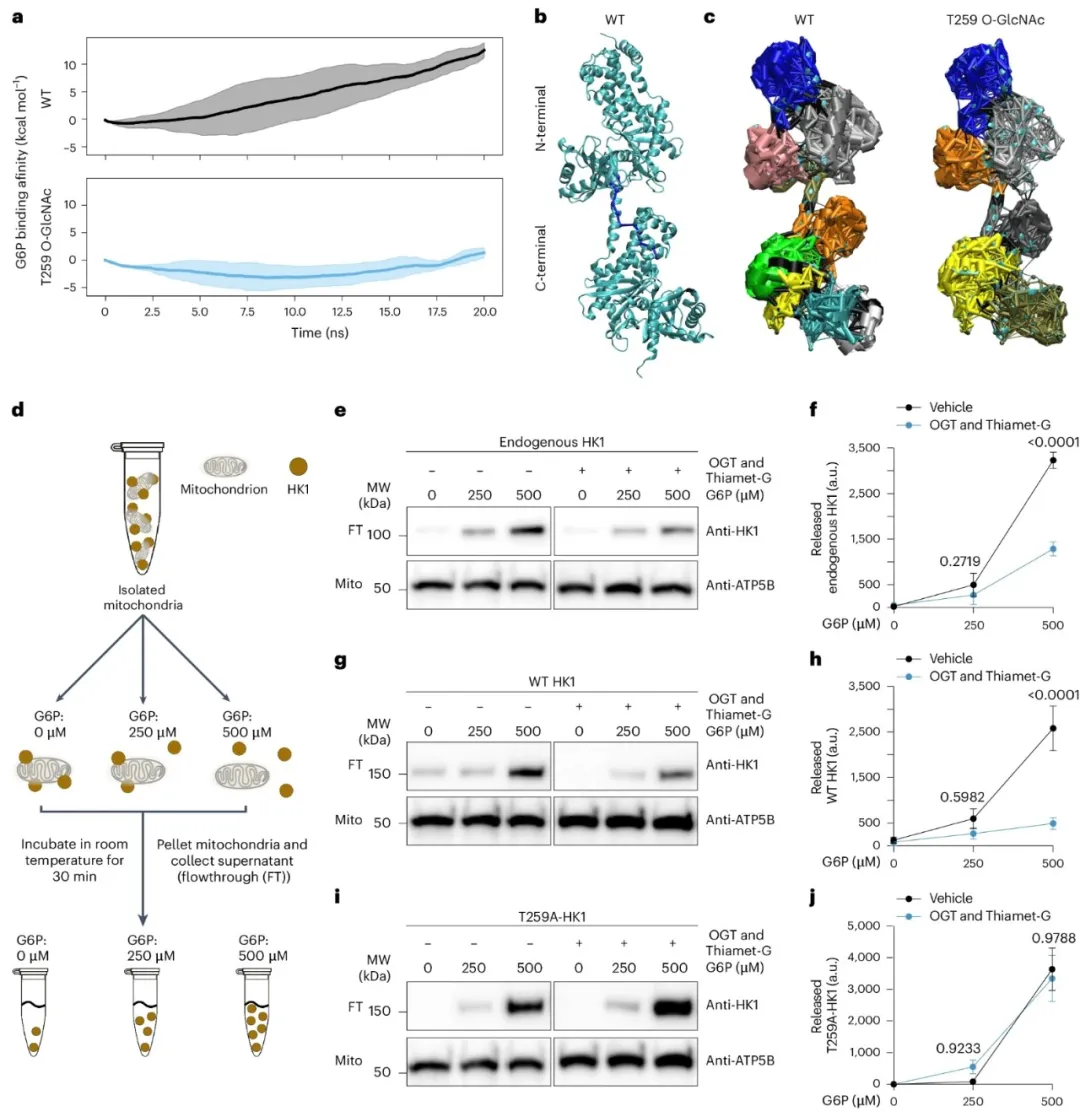
图6. O-GlcNAcylation修饰G6P亲和力并稳定线粒体上的HK1
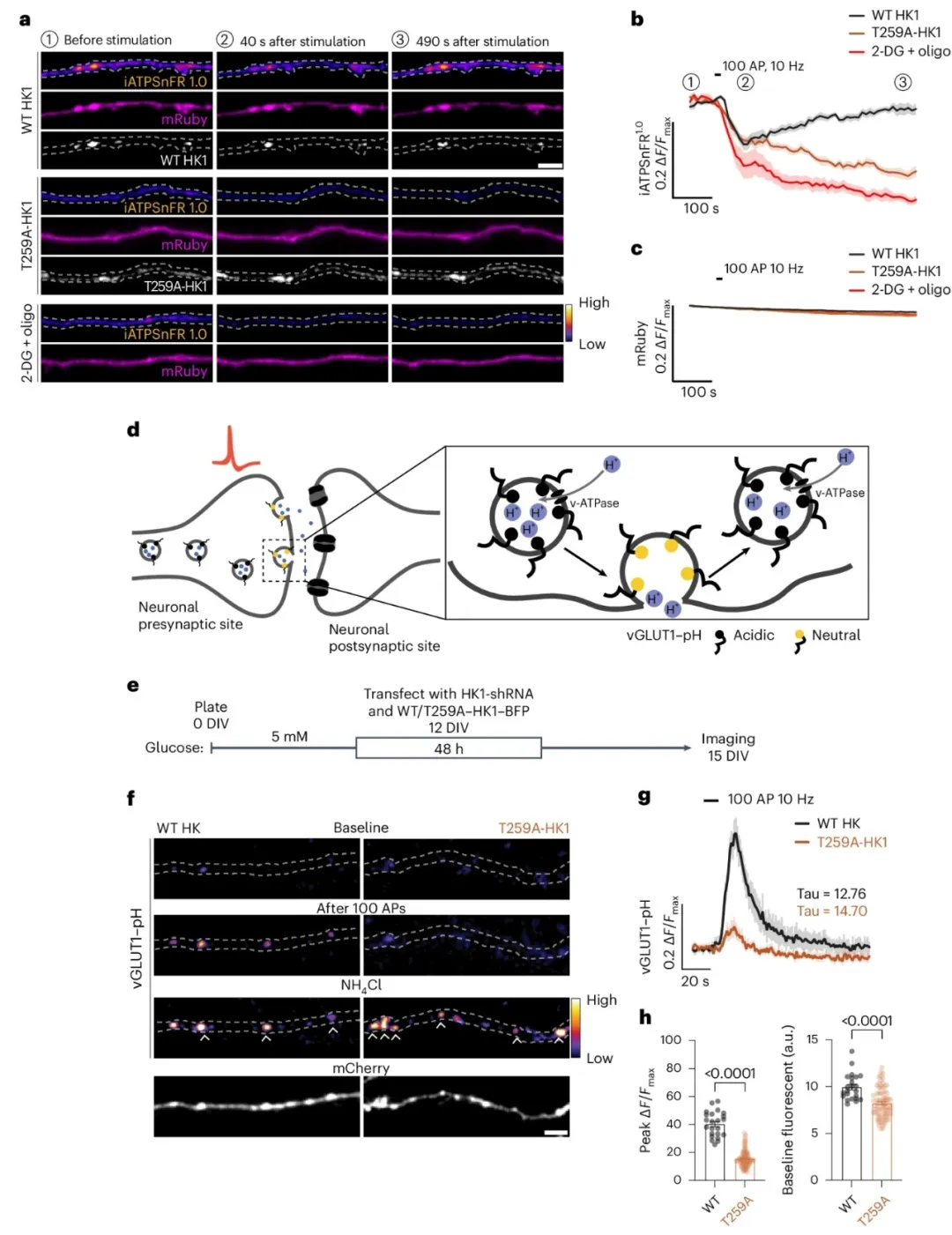
图7. 突触前功能依赖于HK1 O-GlcNAcylation
结论与讨论
本文揭示了HK1的O-GlcNAcylation修饰通过促进其在线粒体上的定位,增强了糖酵解和线粒体ATP产生,从而提高了代谢效率。研究发现,OGT介导的HK1 O-GlcNAcylation不仅调节了HK1的活性,还促进了糖酵解酶的聚集形成代谢体,这对于维持细胞能量平衡和神经功能至关重要。此外,HK1的O-GlcNAcylation对于神经元突触前功能和神经网络活动也起着支持作用。
未来的研究可以深入研究代谢体的组装机制,以及如何通过调节O-GlcNAcylation来优化细胞代谢和治疗相关疾病。未来的工作还可以包括开发针对OGT和OGA的小分子调节剂,以作为潜在的治疗手段,以及利用遗传和药理学工具进一步验证HK1 O-GlcNAcylation在细胞代谢中的功能。